4.1 木材の糖化
4.1.1 パルプ糖化と木材糖化の違い
純粋セルロースであるパルプの糖化と木材の糖化を比較して、木材糖化の特徴を検討した。
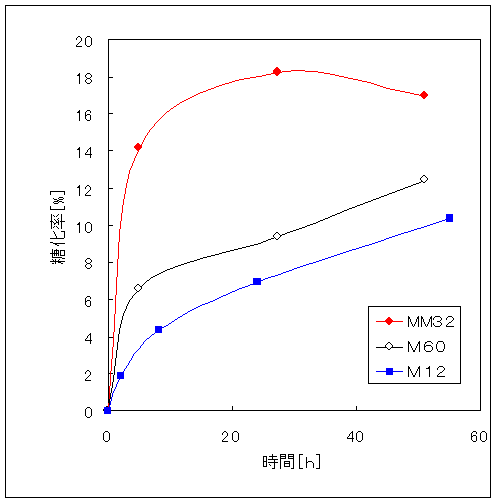
Fig.1はパルプ糖化と木材糖化の実験結果である。比較試料としたパルプは高度に精製された「ろ紙(アドバンテック東洋(株)製の定性分析用濾紙№1)」である。木材にはヒラタケ栽培用の未使用培地(広葉樹を粉砕したもの)をもちいた。一般に木材はセルロース40%、ヘミセルロース30%、リグニン30%からなるリグノセルロース構造をとっている。木材中セルロース含有率40%であることを考慮しても、木材は非常に糖化しにくいことが分かった。文献によると、ろ紙に比べ木材が糖化しにくいのはリグニン、ヘミセルロースの影響によるものである。また、パルプ糖化における糖化速度が8時間前後から大きく減速しているのは、生産された糖による糖化障害(最終産物阻害)と考えられる17,18)。木材の糖化速度も7、8時間前後から大きく減速した。
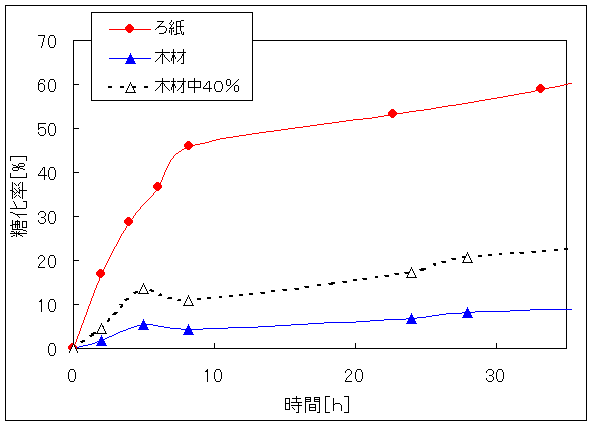
Fig.1 セルラーゼによるパルプ糖化と木材糖化
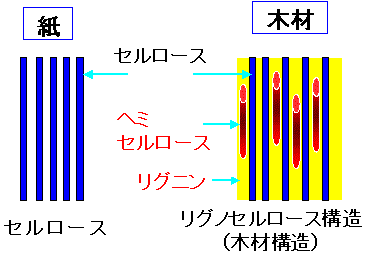
紙と木材構造の違い
4.1.2 酵素濃度が糖化速度に与える影響
Fig.2、Fig.3は木材糖化、パルプ糖化において酵素濃度を変えた場合の糖化率と糖化時間の関係である。この結果から酵素濃度(対試料比)12%よりは36%、60%の方が糖化速度が速い。しかし、酵素濃度に比例した糖化速度の上昇はみられない。酵素濃度36%と60%間では糖化速度と酵素濃度の関係がはっきりしない。また、低濃度、高濃度にかかわらず糖化速度が低下する時間は一致している。以上の結果から、セルラーゼによる糖化には適切な酵素濃度を選択する必要がある。本研究ではこの結果を参考に酵素濃度を12%とした。
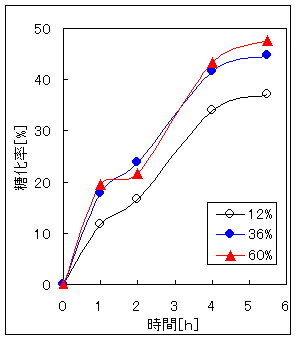
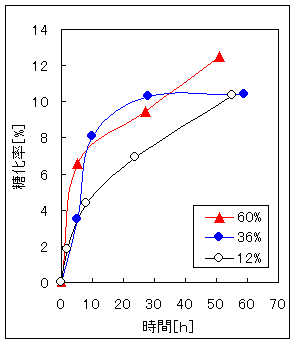
Fig.2 酵素濃度の異なる木材糖化 Fig.3 酵素濃度の異なるパルプ糖化
4.2 ヒラタケ栽培廃棄物の糖化
Fig.4はヒラタケ栽培廃棄物の糖化率と糖化時間の関係である。ヒラタケ1回栽培廃棄物、2回栽培廃棄物、未使用培地の糖化速度には、大きな違いは見られなかった。しかし、栽培培地を熟成させた場合は糖化速度が高まっている。これは、熟成中に菌による木材への作用が進むためであると考えられる。実際、廃棄後一時保管されることから、本研究では熟成した培地についての糖化を検討することとした。
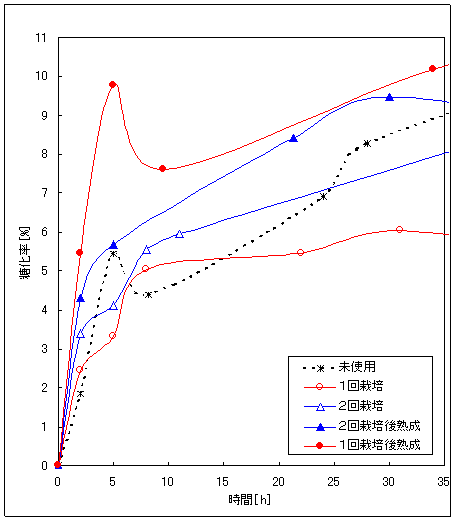
Fig.4 ヒラタケ栽培廃棄物の糖化率と糖化時間
4.3 マイタケ栽培廃棄物の糖化
Fig.5はヒラタケ栽培廃棄物とマイタケ栽培廃棄物の糖化速度を比較したものである。
事前の予想に反して、ヒラタケ栽培廃棄物よりもマイタケ栽培廃棄物の糖化速度がかなり速い結果となった。しかし、マイタケ栽培培地(商業生産品)とヒラタケ栽培培地(自家製)は異なる組成であることを考慮して、さらに検討を進めた。

Fig.5 ヒラタケ栽培廃棄物とマイタケ栽培廃棄物の糖化率と糖化時間


4.4 粒子状態と糖化率
一般に酵素加水分解は酵素と試料の接触面積を大きくすれば糖化速度は高まる14)。そこで、ミルで粉砕したもの(表面積大)と粉砕処理しないもの(表面積小)との糖化速度を比較した。Fig.6はその結果である。木材では糖化速度に変化がなかったが、ヒラタケ栽培廃棄物、マイタケ栽培廃棄物では糖化速度が速くなった。どの程度速くなったかを比較するため粉砕処理試料の糖化率を未処理試料の糖化率で割った粉砕処理変化率と糖化時間の関係(粉砕処理効果)をFig.7に示す。キノコ栽培廃棄物には粉砕処理効果があらわれ、未使用の木材には粉砕処理効果があらわれない。このことから、木材がもともと糖化されにくい構造であり、キノコ栽培廃棄物は糖化されやすい部分の容量が多いことを示している。この点を糖化後の残存物で確認するため、顕微鏡による観察をおこなった。Fig.8は、未糖化の木材と各試料を50時間以上糖化した後の残存物(未反応物)の顕微鏡写真である。ヒラタケ栽培廃棄物よりもマイタケ栽培廃棄物の方が欠損部が多いことから、ヒラタケ栽培廃棄物よりもマイタケ栽培廃棄物の方が糖化されやすい構造であるといえる。
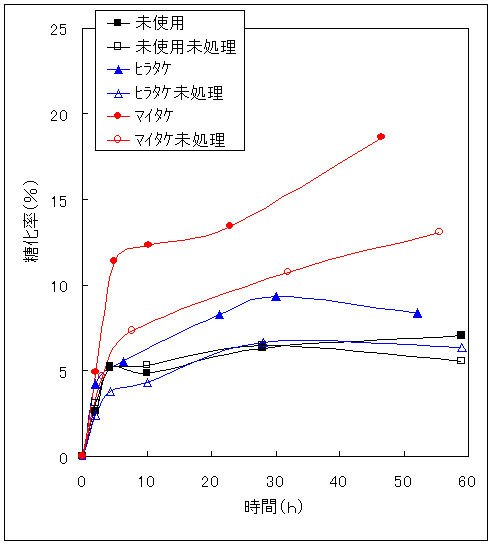
Fig.6 粒子状態と糖化速度
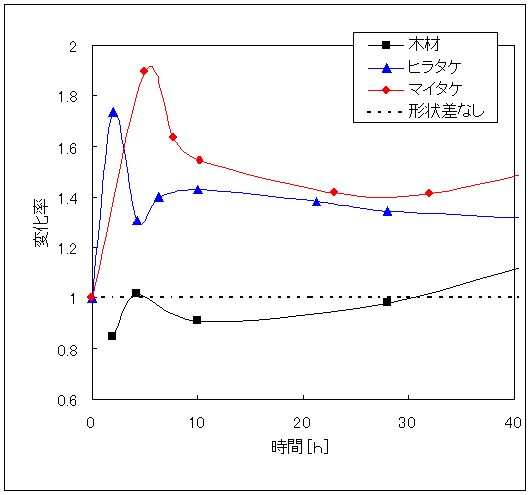
Fig.7 粉砕処理効果(糖化率の変化率)
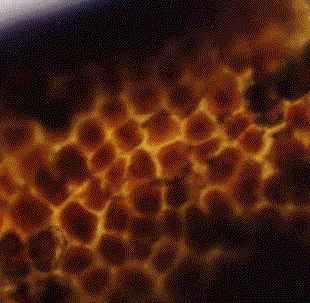
未使用未糖化の木材


ヒラタケ栽培廃棄物 マイタケ栽培廃棄物
Fig.8 未使用未糖化の木材と糖化後の各廃棄物(×100)
4.5 廃棄物中のリグニン量(透過率)
Fig.10は、未使用のマイタケ栽培培地とマイタケ栽培廃棄物である。マイタケ栽培廃棄物は白色化していることから、リグニンが減少していると考えられる。Fig.9は、同じ木材の培地で栽培したヒラタケ栽培廃棄物とマイタケ栽培廃棄物を酢酸中1時間煮沸することで酢酸にリグニンを抽出させた後、その酢酸溶液の透過率を測定したものである。リグニンの溶出しやすさ、リグニン量の減少はともに木材構造の崩壊を示すものである。「4.4粒子状態と糖化率」での考察と総合して検討すると、未使用の木材からの抽出が少ないのは木材構造(リグノセルロース構造)がしっかりしているためであり、ヒラタケ栽培廃棄物からの抽出量が多いことは木材構造が崩れていることを示している。マイタケ栽培廃棄物からの抽出が少ないのは木材構造がしっかりしているためでもあり、木材構造が崩れている部分にリグニンが残っていないためでもある。
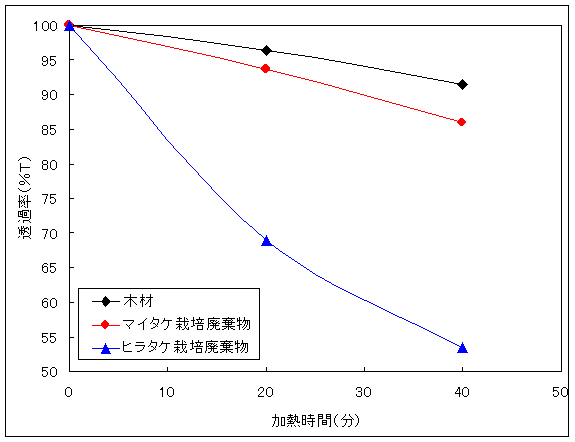
Fig.9 試料を煮込んだ酢酸溶液の透過率